

Cerebelo
Informações de fundo
Crianças SOS têm produzido uma seleção de artigos da Wikipedia para escolas desde 2005. Antes de decidir sobre o patrocínio de uma criança, por que não aprender sobre as diferentes instituições de caridade de patrocínio primeiro ?
| Cérebro: Cerebelo | ||
|---|---|---|
 | ||
| Figura 1-A: Um cérebro humano, com o cerebelo em roxo. | ||
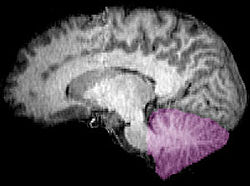 | ||
| Figura 1b: Imagem de ressonância magnética mostrando um médio visão sagital do cérebro humano, com o cerebelo em roxo. | ||
| Parte de | Cérebro | |
| Artéria | SCA, AICA, PICA | |
| Veia | superior, inferior | |
O cerebelo é uma região do cérebro que desempenha um importante papel na integração de percepção sensorial, coordenação e controle motor. A fim de coordenar o controlo do motor, existem muitos vias neurais que ligam o cerebelo com o cerebral córtex motor (que envia informações para o músculos causando-lhes para mover) ea trato espinocerebelar (que fornece feedback proprioceptivo sobre a posição do corpo no espaço). O cerebelo integra estas vias, como um condutor de trem, usando o feedback constante sobre a posição do corpo para movimentos motores afinar.
Devido a esta função de "actualização" do cerebelo, lesões dentro dela não são tão debilitante que cause paralisia, mas sim como presente déficits de feedback, resultando em distúrbios no movimento fino, equilíbrio, postura, e aprendizagem motora. As observações iniciais por fisiologistas durante o século 18 indicam que pacientes com dano cerebelar mostrar problemas com coordenação motora e do movimento. Pesquisa em função cerebelar durante a primeira metade do século 19 foi feita através de estudos de lesões e de ablação em animais . Fisiologistas pesquisa observou que tais lesões levou a animais com movimentos estranhos, andar desajeitado e fraqueza muscular. Estas observações e estudos levaram à conclusão de que o cerebelo foi uma estrutura de controlo do motor. No entanto, a investigação moderna mostra que o cerebelo possui um papel mais amplo em um número de funções cognitivas fundamentais, incluindo a atenção eo processamento de linguagem , música , e outros estímulos sensoriais temporais.
Características gerais
O cerebelo é localizado na porção posterior inferior da cabeça (a rombencéfalo), directamente para o dorsal pons, e inferior ao lóbulo occipital (Figs. 1 e 3). Por causa de seu grande número de pequenas células de grânulos, o cerebelo contém mais do que 50% de todos neurônios no cérebro, mas ocupa apenas 10% do volume total do cérebro. O cerebelo recebe quase 200 milhões de fibras de entrada; em contraste, o nervo óptico é composto por um mero um milhão de fibras.
O cerebelo é dividido em dois grandes hemisférios , muito parecido com o cérebro, e contém dez lóbulos menores. O citoarquitetura ( celular organização) do cerebelo é altamente uniforme, com conexões organizados em um áspero, tridimensional matriz da perpendicular elementos do circuito. Essa uniformidade organizacional faz com que o circuito de nervo relativamente fácil de estudar. Para visualizar esta "matriz perpendicular", alguém poderia imaginar uma rua arborizada com fios correndo em linha reta através dos ramos de uma árvore para outra.
Desenvolvimento e evolução

Durante as fases iniciais de o desenvolvimento embrionário, o cérebro começa a formar-se em três segmentos distintos: o prosencéfalo, mesencéfalo, e rhombencephalon. O rhombencephalon é o mais caudal (para a cauda) segmento do cérebro embrionário; é a partir deste segmento que o cerebelo se desenvolve. Ao longo do segmento rhombencephalic embrionário desenvolver oito inchaços, chamado rhombomeres. O cerebelo surge de duas rhombomeres localizados no placa alar do tubo neural, uma estrutura que, eventualmente, forma do cérebro e da medula espinhal. Os rhombomeres específicos a partir dos quais as formas são cerebelo rhombomere 1 (Rh.1) caudalmente (perto da cauda) e a "istmo" rostral (perto da frente).
Duas regiões primárias são pensados para dar origem aos neurónios que compõem o cerebelo. A primeira zona é a zona ventricular no telhado do quarto ventrículo. Esta área produz Células de Purkinje do cerebelo e profunda neurônios nucleares. Estas células são os neurônios de saída primárias do córtex cerebelar e cerebelo. A segunda zona germinal (berço celular) é conhecido como o lábio rômbico, em seguida, passar por neurónios embrionários semana 27 para a camada granular externa. Esta camada de células encontrados-no exterior do cerebelo-produz os neurônios granulares. Os neurónios granulares migrar a partir desta camada exterior para formar uma camada interna conhecida como a camada interna do grânulo. A camada granular externa deixa de existir no cerebelo madura, deixando apenas células de grânulos na camada interna de grânulos. O cerebelar matéria branca pode ser uma terceira zona germinal no cerebelo; no entanto, a sua função como uma zona germinal é controversa.
O cerebelo é de archipalliar origem filogenética. O palio é um termo para a substância cinzenta que se forma o córtex. O arquipálio é uma das mais evolutivamente regiões cerebrais primitivas. Os circuitos no córtex cerebelar semelhante em todos os classes de vertebrados , incluindo peixes , répteis , pássaros e mamíferos (por exemplo, Fig. 2). Isso tem sido tomado como evidência de que o cerebelo desempenha funções importantes para todos os vertebrados espécies .
Anatomia
O cerebelo contém semelhante cinza e branco divisões importa como o cérebro. Incorporado no interior da matéria branca, que é conhecido como o vitae do mandril (Tree of Life) no cerebelo devido a suas ramificados, treelike aparência-se quatro núcleos cerebelares profundos. Três segmentos filogenéticas brutas são em grande parte agrupados por função geral. As três camadas corticais conter vários tipos celulares que muitas vezes criam vários loops de feedback e feedforward. oxigenado sangue é fornecido por três ramos arteriais ao largo da basilar e artérias vertebrais.
Divisões
O cerebelo podem ser divididos de acordo com três critérios diferentes: anatômico bruto, phyologenetical, e funcionais.
Divisões anatômicas brutas
Por inspecção bruto, três lóbulos pode ser distinguida no cerebelo: o lóbulo flocculonodular, o lobo anterior (rostral à "fissura primária") e o lobo posterior (dorsal para a "fissura primária"). As duas últimas podem ser ainda divididas em uma linha mediana verme cerebelar e hemisférios do cerebelo lateral.
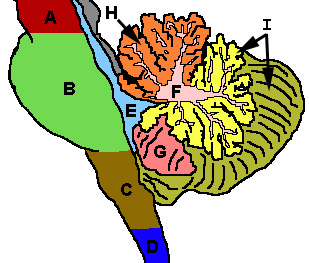 |  |
Divisões filogenéticas e funcionais
O cerebelo também pode ser dividida em três partes com base em ambos critérios filogenéticos (idade evolutiva de cada parte) e sobre os critérios funcionais (as conexões de entrada e saída de cada parte e tem o papel desempenhado em função cerebelar normal). Desde o mais antigo filogeneticamente para os mais novos, as três partes são:
| Denominação funcional (denominação filogenética) | Peças anatômicas | Papel |
| Vestibulocerebelo (Archicerebellum) | Lóbulo Flocculonodular (e vermis imediatamente adjacentes) | O vestibulocerebellum regula movimentos de equilíbrio e de olho. Ele recebe entrada vestibular, tanto do canais semicirculares e da núcleos vestibulares e envia fibras de volta para a medial e núcleos vestibulares laterais. Também recebe input visual do colículos superiores e a partir da córtex visual (esta última através da núcleos pontinos, formando uma via-córtico-ponto cerebelar). As lesões da causa vestibulocerebellum distúrbios de equilíbrio e marcha. |
| Spinocerebellum (Paleocerebellum) | Vermis e partes intermediárias dos hemisférios ("paravermis") | O spinocerebellum regula os movimentos do corpo e dos membros. Ele recebe propriocepção de entrada a partir das colunas dorsais da medula espinal (incluindo o trato espinocerebelar), bem como a partir da do nervo trigeminal, bem como a partir visual e sistemas auditivo. Ele envia fibras para núcleos profundos do cerebelo que por sua vez projecto tanto para o córtex cerebral e o tronco cerebral, proporcionando assim uma modulação de sistemas de motor descendente. O spinocerebellum contém mapas sensoriais que recebe dados sobre a posição de várias partes do corpo no espaço: em particular, as fibras vermis recebe a partir do tronco e as porções proximais dos membros, enquanto que as partes intermédias dos hemisférios recebem fibras a partir das porções distais dos membros . O spinocerebellum é capaz de elaborar entrada proprioceptiva, a fim de antecipar a posição futura de uma parte do corpo durante o curso de um movimento, de uma forma "feed forward". |
| Cerebrocerebellum (Neocerebellum, Pontocerebellum) | Partes laterais do hemisférios | O neocerebellum está envolvido no planejamento de movimento e avaliar a informação sensorial para a ação. Ele recebe entrada exclusivamente a partir do córtex cerebral (especialmente o lobo parietal) através do núcleo pontino (formando caminhos-cortico-cerebelar Ponto), e envia fibras principalmente para o ventrolateral tálamo (por sua vez ligado a áreas motoras do córtex pré-motor e área do motor primário do córtex cerebral) e para o núcleo vermelho (por sua vez ligado ao núcleo olivar inferior, que liga de volta para os hemisférios do cerebelo). O neocerebellum está envolvido em movimento planejamento que está prestes a ocorrer e tem funções puramente cognitivos também. |
Muito do que se sabe sobre as funções do cerebelo decorre da documentação cuidadosa dos efeitos de lesões focais em pacientes humanos que sofreram lesão ou doença ou lesão por meio de pesquisa animal.
Núcleos profundos
Os núcleos profundos do cerebelo ato como os principais centros de comunicação, e os quatro núcleos diferentes do cerebelo (denteado, interpositus, fastigial e vestibular) receber e enviar informações para partes específicas do cérebro. Além disso, estes núcleos receber ambos os sinais inibitórios e excitatórios a partir de outras partes do cérebro que por sua vez afectam os sinais de saída do núcleo.
Camadas corticais


Existem três camadas para o córtex cerebelar; do invólucro exterior para a camada interior, estas são as camadas moleculares, Purkinje e granular. A função do córtex cerebelar é, essencialmente, para modular a informação que flui através dos núcleos profundos. O microcircuito do cerebelo é esquematizada na Figura 5. Mossy e fibras de escalada transportar informações sensório-motor nos núcleos profundos, que por sua vez passá-lo para diversas áreas pré-motoras, regulando assim a ganhar eo calendário das ações motoras. Mossy e fibras de escalada também se alimentam esta informação no córtex cerebelar, que executa vários cálculos, resultando na regulação de Purkinje disparo das células. Neurônios de Purkinje realimentam os núcleos profundos através de um potente inibidor sinapse. Este sinapse regula o grau em que as fibras musgosas e escalada activar os núcleos profundos, e, assim, controlar o efeito final do cerebelo em função motora. A força sináptica de quase todos os sinapse no córtex cerebelar foi mostrado para sofrer plasticidade sináptica. Isto permite que o circuito do córtex cerebelar para ajustar continuamente e ajustar a saída do cerebelo, formando a base de certos tipos de aprendizagem e coordenação motora. Cada camada no córtex cerebelar contém os vários tipos de células que compõem este circuito.
Camada granular
A camada mais interna contém os corpos celulares dos dois tipos de células: a numerosos e minúsculos células de grânulos, e a maior Células de Golgi. Fibras Mossy entra a camada granular do seu principal ponto de origem, os núcleos pontinos. Estas fibras formam sinapses excitatórias com as células granulares e as células dos núcleos profundos do cerebelo. As células granulares enviar seus axônios conhecido em forma de T como paralelo fibras-se para dentro da camada superficial molecular, onde formam centenas de milhares de sinapses com células de Purkinje dendritos. O cerebelo humano contém na ordem de 60 a 80 mil milhões de células granulares, tornando este tipo de célula única, de longe, os mais numerosos neurónios no cérebro (cerca de 70% de todos os neurónios no cérebro e na medula espinhal, combinado). Células de Golgi fornecer feedback inibitório para células de grânulos, formando com eles uma sinapse e que se projecta de um axónio na camada molecular.
Camada de Purkinje
A camada intermédia contém somente um tipo de corpo da célula, isto o elevado Células de Purkinje. Células de Purkinje são os neurônios de integração primários do córtex cerebelar e fornecer seu exclusivo saída. Dendrites Purkinje são grandes mandris com centenas de galhos espinhosos atingindo-se na camada molecular (Fig. 6). Estes mandris dendríticas são flat-quase todos eles se encontram em planos-com a vizinha mandris de Purkinje em planos paralelos. Cada fibra paralelo das células de grânulos é executado ortogonal através destes mandris, como um fio passando por muitas camadas. Neurônios de Purkinje são GABAergic-o que significa que têm sinapses inibidoras-com os neurônios do cerebelo profunda e núcleos vestibulares no tronco cerebral. Cada célula Purkinje recebe a entrada excitatória de 100.000 para 200.000 fibras paralelas. Fibras paralelas são referidos como sendo responsável pela simples (tudo ou nada, amplitude invariante) spiking da célula de Purkinje.
Células de Purkinje também receber o contributo do núcleo olivar inferior via fibras de escalada. Um bom mnemônico para essa interação é a frase "subir a outra oliveira", dado que as fibras de escalada originam da oliva inferior contralateral. Em flagrante contraste com as 100.000-plus entradas de fibras paralelas, cada uma das células de Purkinje recebe entrada de fibra exatamente uma escalada; mas esta única fibra "sobe" os dendritos das células de Purkinje o, enrolando em torno deles e fazer um grande número de sinapses em que vai. A entrada de líquido é tão forte que uma única acção potencial a partir de uma fibra de escalada é capaz de produzir um "pico complexo" na célula de Purkinje: um rebentamento de vários picos de uma linha, com amplitudes decrescentes, seguidas por uma pausa, durante o qual picos simples são suprimidas.
Camada Molecular
Esta camada mais externa do córtex cerebelar contém dois tipos de inibidor interneurónios: o estreladas e células cesta. Ele também contém os mandris dendríticas de neurônios de Purkinje e feixes de fibras paralelas a partir de células granulares. Ambas as células estreladas e cesta formar sinapses GABAérgicos Onto dendrites de Purkinje.
Pedúnculos
Da mesma forma, o cerebelo segue a tendência de "trios", com três grandes pedúnculos de entrada e saída (feixes de fibras). Estes são o superior (conjunctivum brachium), médio (pontis brachium) e inferior (corpo restiform) pedúnculos cerebelares.
| Pedúnculo | Descrição |
| Superior | Embora existam algumas fibras aferentes do trato espinocerebelar anterior que são transportadas para o lóbulo anterior do cerebelo através deste pedúnculo, a maioria das fibras são fibras eferentes. Assim, o pedúnculo cerebelar superior é a principal via de saída do cerebelo. A maioria das fibras eferentes originam dentro do núcleo denteado que por sua vez projeto a vários estruturas mesencéfalo incluindo o núcleo vermelho, o núcleo lateral ventral / ventral anterior da tálamo, eo medula. O dentatorubrothalamocortical (denteado núcleo> núcleo vermelho> tálamo> córtex pré-motor) e cerebellothalamocortical (cerebelo> tálamo> córtex pré-motor) Vias são duas vias principais que passam por este pedúnculo e são importantes no planejamento motor. |
| Meio | Este é composto inteiramente de fibras aferentes com origem na núcleos pontinos como parte do maciço trato corticopontocerebellar (córtex cerebral> pons> cerebelo). Estas fibras descem das áreas sensoriais e motoras do cerebral neocortex e fazer o pedúnculo cerebelar médio o maior dos três pedúnculos cerebelares. |
| Inferior | Esta carrega muitos tipos de fibras de entrada e saída que estão principalmente preocupados com a integração input sensorial proprioceptiva com motor funções vestibulares, como equilíbrio e manutenção da postura. Informações proprioceptivas do corpo é transportado para o cerebelo através dorsal trato espinocerebelar. Este aparelho passa através do pedúnculo cerebelar inferior e sinapses dentro do paleocerebellum. Vestibular informações projecta no archicerebellum. O escalada fibras do oliva prazo inferior através do pedúnculo cerebelar inferior. Este pedúnculo também traz informações diretamente do Células de Purkinje para o núcleos vestibulares no tronco cerebral dorsal localizado na junção entre a pons e medula. |
Existem três fontes de entrada para o cerebelo, em duas categorias constituídos de fibras cobertas de musgo e escalada, respectivamente. Mossy fibras podem se originar a partir dos núcleos pontinos, que são grupos de neurônios localizados na ponte que carregam a informação a partir do córtex cerebral contralateral. Eles também podem surgir dentro do trato espinocerebelar cuja origem está localizada na ipsilateral medula espinhal. A maior parte da saída do cerebelo sinapses inicialmente sobre os núcleos profundos do cerebelo, antes de sair através das três pedúnculos. A exceção mais notável é a inibição direta dos núcleos vestibulares por células de Purkinje.
Fornecimento de sangue

Três artérias fornecem sangue para o cerebelo (Fig. 7): o artéria cerebelar superior (SCA), artéria cerebelar inferior anterior (AICA), e artéria cerebelar inferior posterior (PICA).
Artéria cerebelar superior
Os ramos SCA fora da porção lateral da artéria basilar, apenas inferior à sua bifurcação para a artéria cerebral posterior. Aqui ela envolve posteriormente em torno da ponte (a que também fornece sangue) antes de chegar ao cerebelo. A SCA fornece sangue para a maior parte do córtex cerebelar, os núcleos do cerebelo, e os pedúnculos cerebelares médios e superiores.
Artéria cerebelar inferior anterior
Os ramos AICA fora da porção lateral da artéria basilar, apenas superior à junção das artérias vertebrais. Desde a sua origem, se ramifica ao longo da parte inferior da ponte no ângulo pontocerebelar antes de atingir o cerebelo. Esta artéria fornece sangue para a porção anterior do cerebelo inferior, e para o facial (NC VII) e nervos vestibulococleares (NC VIII).
Obstrução da AICA pode causar paresia, paralisia e perda de sensibilidade na face; ela também pode causar deficiência auditiva . Além disso, poderia causar um infarto do ângulo pontocerebelar. Isto pode levar a hiperacusia (disfunção do músculo estapédio, inervado por NC VII) e vertigens (interpretação errada do vestibular canal semi-circular de aceleração da endolinfa provocado pela alteração de CN VIII).
Artéria cerebelar inferior posterior
Os ramos PICA fora da porção lateral das artérias vertebrais apenas inferior à sua junção com a artéria basilar. Antes de atingir a superfície inferior do cerebelo, o PICA envia ramos dentro da medula, o fornecimento de sangue a vários núcleos dos nervos cranianos. No cerebelo, a PICA fornece o sangue à porção posterior inferior do cerebelo, no pedúnculo cerebelar inferior, o núcleo ambíguo, a núcleo motor do vago, a medula núcleo trigeminal, a núcleo solitário, eo núcleos vestibulococleares.
Função Geral
Funcionalmente, o fibra e a escalada caminhos-de fibras musgosas grânulo de células-paralelo de fibras são os dois principais tipos de aferentes para o cerebelo como um todo e para as células de Purkinje, em particular. Estes sistemas aferentes diferir dramaticamente na sua conectividade. A célula de Purkinje e suas vias aferentes fibra de escalada tem um relacionamento um-para-um e de projeção global é organizada para produzir ativação síncrona de grupos específicos de células de Purkinje em uma orientação rostrocaudal. A relação entre a célula de Purkinje e o sistema de fibras musgosas de fibra paralela pode ser caracterizada como de muitos para muitos. Com a direccionalidade ser orientação mediolateral dentro da camada molecular, ou seja, em ângulo recto com os dendritos de células de Purkinje, que são isoplanar.
O sistema de fibra de escalada
Origina a partir da oliva inferior contralateral. Como resultado do acoplamento eléctrico entre neurónios oliva inferior, o seu desacoplamento dinâmico através da inibição de retorno a partir dos núcleos cerebelares e a topografia da projecção olivocerebellar, este sistema gera síncrono (numa escala de tempo de milisegundos) activação pico complexo de células de Purkinje, em rostrocaudally bandas orientadas. Estas bandas de atividade são cerca de 250 m de largura na direção médio-lateral, mas pode ser de vários milímetros de comprimento na direção rostrocaudal e estender para baixo as paredes da folia cerebelar e em toda a vários lóbulos. A distribuição sincronia momento-a-momento de controle motor é dinamicamente modulado pelo oliva inferior com o importante papel das aferentes olivary sendo para determinar o padrão de acoplamento eletrônico "eficaz" entre os neurônios olivary e assim a distribuição de pico de atividade complexa síncrona através o córtex cerebelar. Alterações nos padrões de sincronia estão associados a movimentos feitos pelos animais que executam uma tarefa motora .. De fato. O sistema olivocerebellar pode ser considerado um substrato electricamente maleável a partir do qual as sinergias único motor pode ser esculpido.
O sistema de fibra Mossy Fiber-Parallel
Em contraste com a natureza ponteada de activação cerebelar pelo sistema olivocerebellar, o sistema de fibras musgosas-fibra paralelo fornece uma regulação contínua e muito delicado da excitabilidade dos núcleos cerebelares , provocada pela activação tónico de picos simples em células de Purkinje, que em última instância, gera o controle fino do movimento conhecido como coordenação motora. O facto de as fibras musgosas informar o córtex cerebelar tanto ascendente e descendente mensagens de e para os centros motores na medula espinhal e do tronco cerebral nos dá uma idéia do papel final do sistema de fibras musgosas: informa o córtex do lugar e taxa de movimento dos membros e coloca as intenções do motor gerados pelo cérebro no contexto do estado do corpo no momento do movimento é para ser executado. Além disso, através de seus efeitos sobre as células nucleares inibitórios GABAérgicos do cerebelo, que projetam de volta para a oliva inferior, que ajuda a moldar o padrão de acoplamento entre as células olivary e, consequentemente, a distribuição sincronia na descarga olivocerebellar próximo.
O cerebelar Núcleos
As células de Purkinje são a única saída do córtex cerebelar e são inibitórios na natureza Seus axônios em contato com o cerebelo e Deiters núcleo vestibular como seu único alvo. A actividade dos núcleos cerebelares é regulada em três formas: (1), através de estímulos excitatórios a partir colaterais dos sistemas de aferentes cerebelares, (2) através de entradas de inibição de células de Purkinje activadas através das vias das fibras musgosas, e (3) através de entradas de Purkinje células activadas por o sistema de fibra de escalada
Função cerebelar geral
A saída do cerebelo (os axónios do núcleo cerebelar) siga para gerar a actividade de fundo que serve para ajustar o tom global e postura que dá o córtex motor a capacidade de executar movimentos com base na intenção (a estratégia de movimento). Neste contexto, o cerebelo fornece as táticas da ativação muscular múltipla necessária para suportar tais definir movimentos. E assim, enquanto o cérebro do motor determina para onde mover (imperativo executivo) do cerebelo implementa seu momento adequado e modula a força dada a cada comando do motor, como a coordenação de movimento é uma função não-contínua.
Disfunção
A ataxia é um complexo de sintomas, que envolve geralmente uma falta de coordenação, que é muitas vezes encontrada em processos patológicos que afectam o cerebelo. Para identificar os problemas cerebelares, os exame neurológico inclui a avaliação da marcha (a marcha de base ampla sendo indicativo de ataxia), testes apontando dedo e avaliação de postura. Anormalidades estruturais do cerebelo (hemorragia, infarto, neoplasia, degeneração) podem ser identificados em imagens de corte transversal. A ressonância magnética é a modalidade de escolha, como tomografia computadorizada não é suficientemente sensível para detectar anormalidades estruturais do cerebelo.
Envelhecimento
A estudo estereológico descobriu que cerebelar humano substância branca foi reduzida em 26% com idade (na gama de 19-84 anos). Os investigadores do estudo podem não detectar a perda global de Purkinje ou células de grânulos, no entanto, no lobo anterior houve uma perda significativa destes tipos de células, bem como uma perda de volume de 30%. Com Imagem por Ressonância Magnética uma redução volumétrica moderada com a idade em vermis e hemisfério cerebelar foi observado.
Um estudo auto-radiografia do cerebelo humano encontrou uma crescente a ligação de H-3- cetanserina com a idade. (Cetanserina se liga principalmente à 5-HT 2A neuroreceptor) A mesma equipa de investigação não encontrou nenhuma correlação significativa com a idade em sua homogeneizado estudo de ligação. Um pouco em conformidade com o estudo de uma autorradiografia estudo por tomografia de emissão de pósitrons com o altanserina do receptor 5-HT2A radioligante encontrou uma correlação positiva entre a idade ea cerebelar ligação não específica.
Teorias sobre a função cerebelar
Duas teorias principais abordar a função do cerebelo, tanto lidar com a coordenação motora. Uma afirma que as funções do cerebelo como um regulador do "calendário de movimentos". Este surgiu a partir de estudos de pacientes cujos movimentos cronometrados são interrompidas.
O segundo, "Teoria da Rede Tensor" fornece um modelo matemático de transformação da sensorial (covariante) coordenadas espaço-temporais em motor (contravariante) coordena por redes neuronais do cerebelo.
Como muitas controvérsias nas ciências físicas, não há evidências que suportam cada uma das hipóteses acima. Estudos de aprendizagem motora no reflexo vestíbulo-ocular e condicionamento eyeblink demonstrar que o calendário e amplitude de movimentos aprendidas são codificados pelo cerebelo. Muitos mecanismos de plasticidade sináptica foram encontrados em todo o cerebelo. O modelo de Marr-Albus atribui principalmente a aprendizagem motora a um único mecanismo de plasticidade: o depressão a longo prazo das sinapses de fibras paralelas. A Teoria da Tensor Rede de transformações sensório-motores por o cerebelo também tem sido experimentalmente suportado.
Com o advento de mais sofisticado técnicas de neuroimagiologia, tais como A tomografia de emissão de positrões (PET), e IRMf, numerosos diversas funções estão agora, pelo menos parcialmente atribuída ao cerebelo. O que já foi pensado para ser principalmente uma / região de integração sensorial motor está agora provando ser envolvido em muitos diversas funções cognitivas.
Modelagem cerebelar
Tal como mencionado na secção anterior, tem havido muitas tentativas para modelar a função cerebelar. Os insights fornecidos pelos modelos também levaram a extrapolações nos domínios da metodologias de inteligência artificial, especialmente redes neurais. Algumas das realizações notáveis foram Cerebellatron, Cerebelar modelo associativo de memória ou redes CMAC, e SpikeFORCE para controle de movimento robótico, e "Teoria da Rede Tensor".
Imagens adicionais
- CT do cérebro de Mikael Häggström S3 I8.JPG
A tomografia computadorizada da cabeça, com cerebelo visível em parte inferior.

Lobos

Diencéfalo

Esquema mostrando as ligações das várias partes do cérebro.

A superfície superior do cerebelo.

Sob a superfície do cerebelo.

Corte sagital do cerebelo, perto da junção do verme com o hemisfério.

A dissecção mostrando as fibras de projecção do cerebelo.

Esquema de telhado do quarto ventrículo. A seta é no forame de Majendie.
Dissection que mostra a evoluo das fibras cefalorraquidiano.

Diagrama mostrando as posições das três principais cisternas subaracnóide.

Cerebelo humano visão anterior

Cérebro humano vista sagital médio
