

Theropoda
Você sabia ...
Crianças SOS feita esta seleção Wikipedia ao lado de outras escolas recursos . SOS mães cada um cuidar de uma família de crianças apadrinhadas .
| Terópodes Intervalo temporal: Triássico-presente, 231.4-0Ma | |
|---|---|
 | |
| Réplica montada de um Tyrannosaurus rex esqueleto | |
| Classificação científica | |
| Reino: | Animalia |
| Filo: | Chordata |
| Classe: | Reptilia |
| Ordem: | Saurischia |
| Clado: | Eusaurischia |
| Clado: | Theropoda Marsh, 1881 |
| Subgrupos | |
| |
Theropoda (terópode / θ ɛr ə p ɒ d /; nome subordem Theropoda / θ ɨ r ɒ p ɵ d ə /, Do grego que significa "pés animal") é tanto uma subordem dos bípedes saurischian dinossauros , e um clado consiste em que subordem e seus descendentes (incluindo modernas aves ). Dinossauros pertencentes à subordem Theropoda eram principalmente carnívoros , embora um número de grupos de terópodes evoluiu herbivoria , onívoria, e insetivoria. Terópodes apareceu pela primeira vez durante o Idade Carniano do final do Triássico período de cerca de 230 milhões de anos atrás ( Ma) e incluiu os únicos grandes carnívoros terrestres do Jurássico adiantado até pelo menos o fim do Cretáceo , cerca de 65 Ma. No Jurassic , pássaros evoluíram a partir de pequenos especializado terópodes coelurosaurian, e são hoje representados por 9.900 espécies vivas.
Entre as características que ligam dinossauros terópodes para pássaros são o pé-de-coleira, um fúrcula (wishbone), ossos cheios de ar, ninhada do ovos, e (em alguns casos) penas.
Biologia
Dieta

Terópodes exibem uma ampla gama de dietas, de insetívoros para os herbívoros e carnívoros. Carnivory Strict sempre foi considerado a dieta ancestral para terópodes, como um grupo, e uma ampla variedade de dietas foi historicamente considerada uma característica exclusiva para os terópodes aviária (pássaros). No entanto, as descobertas no final dos anos 20 e início do século 21 mostraram que uma variedade de dietas existiu mesmo em mais linhagens basais. Todos os achados iniciais de fósseis de terópodes mostrou-lhes a ser essencialmente carnívoros . Espécimes fossilizados de terópodes início conhecidos cientistas dos séculos 20 e início dos anos 19 todos possuíam dentes afiados com bordas serrilhadas para cortar carne, e alguns exemplares ainda mostrou evidências diretas de comportamentos predatórios. Por exemplo, um Compsognathus longipes fóssil foi encontrado com um lagarto em seu estômago, e um Velociraptor mongoliensis espécime foi encontrado trancado em combate com um Protoceratops andrewsi (um tipo de ornithischian dinossauro).
Os terópodes fósseis não-carnívoros primeiro confirmadas encontradas foram a therizinosaurs, originalmente conhecida como segnosaurs. Primeiro pensado para ser prossaurópodes, estes dinossauros enigmáticas foram mais tarde provou ser altamente especializados, herbívoros terópodes. Therizinosaurs possuía grandes abdomens para alimento de planta de processamento e cabeças pequenas com bicos e dentes em forma de folha. Um estudo mais aprofundado de terópodes maniraptoran e suas relações mostrou que therizinosaurs não eram os únicos membros iniciais deste grupo a abandonar carnivory. Várias outras linhagens de início maniraptors mostram adaptações para um dieta onívora, incluindo-semente comer (alguns troodontídeos) e inseto-comer (muitos avialans e alvarezssauros). Oviraptorosaurs, ornithomimosaurs e troodontídeos avançados eram susceptíveis onívoro, bem como, e alguns terópodes iniciais (tais como Masiakasaurus knopfleri eo espinossaurídeos) parecem ter-se especializado na captura de peixe.
Pele, escamas e penas
Terópodes Mesozóico foram também muito diversificada em termos de textura da pele e cobertura. Penas ou estruturas pena-como são atestados na maioria linhagens de terópodes. (Ver dinossauro emplumado). No entanto, fora do coelurosaurs, penas podem ter sido confinada às espécies pequenas, menores, ou partes do animal limitado. Muitos terópodes maiores tinham pele coberta em pequenas escalas, esburacadas. Em algumas espécies, estes foram intercaladas com escalas maiores, com núcleos ósseos, ou osteoderms. Este tipo de pele é mais conhecido no Ceratosaur Carnotaurus, que foi preservada com extensas impressões de pele.
A partir de 2012, o terópode protofeathered conhecido mais divergente é Sciurumimus albersdoerferi. Assim, pode-se inferir que Tetanurae foram pelo menos parcialmente de penas; nenhuma evidência de penas (Proto) foi descoberto entre o Ceratosauria. As linhagens mais distantes dos pássaros tinham penas que eram relativamente curto e composto por simples possivelmente ramificação filamentos,. Filamentos simples também são vistos em therizinosaurs, que também possuía grande, endureceu "pena", como penas. Mais plenamente terópodes emplumados, como dromeossauros, normalmente conservam apenas as escalas pés. Algumas espécies podem ter penas mistos noutras partes do corpo também. Epidendrosaurus preservada escalas perto da parte inferior da cauda, e Juravenator pode ter sido predominantemente escamosa com alguns filamentos simples intercaladas. Por outro lado, alguns terópodes foram completamente cobertas com penas, tais como o troodontídeos Anchiornis, que ainda tinha penas nos pés e dedos dos pés.
Tamanho
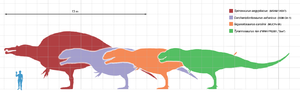
Tiranossauro era o maior e mais popular terópode conhecido do público em geral para muitas décadas. Desde a sua descoberta, no entanto, foram descritos uma série de outros dinossauros carnívoros gigantes, incluindo Espinossauro , Carcharodontosaurus, e Giganotosaurus . Os espécimes originais Spinosaurus (bem como os fósseis mais recentes descritos em 2006) apoiam a ideia de que Spinosaurus é maior do que o tiranossauro, mostrando que Spinosaurus era possivelmente 6 metros a mais e, pelo menos, 1 tonelada mais pesado que o tiranossauro. Não há ainda nenhuma explicação clara para exatamente por que estes animais cresceu muito maior do que os predadores terrestres que vieram antes e depois deles.
A menor terópode não avialan conhecido a partir de espécimes adultos é a troodontídeos Anchiornis huxleyi, em 110 gramas de peso e 34 cm (1 pé) de comprimento. Quando os pássaros modernos são incluídos, o Bee Hummingbird Mellisuga helenae é menor em 1,9 g e 5,5 cm (2,2 pol) de comprimento.
Postura e marcha
Como um grupo extremamente diversificado de animais, a postura adotada por terópodes provável variou consideravelmente entre várias linhagens ao longo do tempo. Todos os terópodes conhecidos são bípede, com os membros anteriores reduzidos em comprimento e especializadas para uma grande variedade de tarefas (ver abaixo). Nas aves modernas, o corpo é tipicamente mantida numa posição um tanto na posição vertical, com a parte superior da perna (fémur), realizada paralela à coluna vertebral e com a força de locomoção para a frente gerado pelo joelho. Os cientistas não têm certeza de quão longe de volta na árvore genealógica terópode este tipo de postura e locomoção estende.
Terópodes não-aviários foram reconhecidos pela primeira vez como bípede durante o século 19, antes de sua relação com aves foi amplamente aceita. Durante este período, terópodes, como carnossauros e tyrannosaurids foram pensados para ter caminhado com fêmures verticais e espinhos em uma posição vertical, quase postura ereta, usando suas caudas longas e musculosas como suporte adicional em uma posição tripodal canguru-like. Começando na década de 1970, estudos biomecânicos de terópodes gigantes extintos em dúvida essa interpretação. Estudos de articulação óssea dos membros e da relativa ausência de provas trackway para a cauda arrastando sugeriram que, ao caminhar, o gigante, terópodes de cauda longa teria adoptado uma postura mais horizontal com a cauda realizada paralela ao solo. No entanto, a orientação das pernas nestas espécies enquanto curta permanece controverso. Alguns estudos apoiar um fêmur de orientação vertical tradicional, pelo menos nas maiores terópodes de cauda longa, enquanto outros sugerem que o joelho estava normalmente fortemente flexionado em todos os terópodes durante a caminhada, até mesmo gigantes como os tyrannosaurids. É provável que uma vasta gama de posturas corporais, posturas e andamentos existia nos diversos grupos terópodes extintos.
Sistema nervoso e os sentidos
Embora, elencos completos raros de terópode endocrania são conhecidos a partir de fósseis. Terópode endocrania também pode ser reconstruído a partir de casos cerebrais preservadas sem danificar as amostras valiosas usando uma tomografia computadorizada e software de reconstrução 3D. Estes achados são de importância evolutiva, porque eles ajudam documentar o surgimento da neurologia das aves modernas do que de répteis anteriores. Um aumento na proporção do cérebro ocupado pelo cérebro parece ter ocorrido com o advento da Coelurosauria e "continua ao longo da evolução maniraptorans e madrugadores ".
Morfologia membro anterior

Forelimbs curtos em relação ao patas traseiras era um traço comum entre os terópodes, principalmente no abelisaurids (tal como Carnotauro) eo tyrannosaurids (como o Tyrannosaurus ). Esta característica foi, no entanto, não é universal: espinossaurídeos tinha bem desenvolvida membros dianteiros, assim também fizeram muitos coelurosaurs. Um gênero, Xuanhanosaurus, foi também alegou ter sido quadrúpede por causa de suas patas dianteiras relativamente robustos, mas isso não é mais pensado para ser provável.
As mãos também são muito diferentes entre os diferentes grupos; o mais comum é um apêndice que consiste em três dedos, os dígitos I, II e III (ou, possivelmente, II, III e IV) com garras afiadas. Alguns terópodes basais ( Herrerasaurus , Eoraptor) teve 4 dígitos, e também uma reduzida V. metacarpo Ceratosaurians geralmente tinham quatro dígitos, enquanto a maioria dos tetanurans teve 3.
Âmbito de utilização dos membros anteriores, também se acredita também ter sido diferente entre as famílias diferentes. O espinossaurídeos poderia ter usado suas patas dianteiras poderosas para segurar peixe. Alguns pequenos coelurosaurus ( dromeossaurídeos, scansoriopterygids) Acredita-se que usaram suas patas dianteiras para subir em árvores e pássaros, que se acredita serem descendentes dos terópodes, usá-los para o vôo.
Movimento forelimb
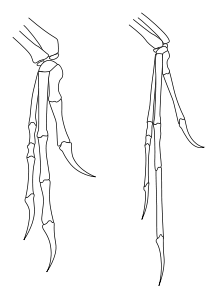
Ao contrário do que a forma como os terópodes têm sido muitas vezes reconstruídos na arte e na mídia popular, a amplitude de movimento das patas dianteiras terópodes era severamente limitada, especialmente em comparação com a destreza dos membros anteriores dos seres humanos e outros primatas. Mais notavelmente, terópodes e outros dinossauros bípedes saurischian (incluindo os bípedes prossaurópodes ) não puderam pronate suas mãos, isto é, eles não poderia rodar o antebraço de modo a que as palmas enfrentou o solo ou para trás, para as pernas. Nos seres humanos, é conseguida através de pronação movimento do relativamente ao raio cúbito (os dois ossos do antebraço). Em dinossauros saurischian, no entanto, a fim de o raio perto do cotovelo foi efectivamente bloqueada numa ranhura do cúbito, impedindo qualquer movimento. Movimento no pulso também foi limitado em muitas espécies, forçando todo o antebraço e mão para se mover como uma unidade única, com pouca flexibilidade. Em terópodes e prossaurópodes, a única maneira para a palma da mão para enfrentar o terreno teria sido por splaying lateral inteira do membro anterior, como em uma ave levantando seu braço.
Em carnossauros como Acrocanthosaurus, a própria mão retido um grau relativamente elevado de flexibilidade, com dedos móveis. Este também foi verdadeiro de mais terópodes basais como herrerasaurs e dilophosaurs. Coelurosaurs mostrou uma mudança na utilização do antebraço, com maior flexibilidade no ombro permitindo que o braço para ser levantado em relação ao plano horizontal, e ainda maiores graus em aves que voam. No entanto, em tais como coelurosaurs ornithomimosaurs e especialmente dromeossauros, a própria mão tinha perdido a maior flexibilidade, com os dedos altamente inflexíveis. Dromeossauros e outros maniraptorans também mostrou aumento da mobilidade no pulso não visto em outros terópodes, graças à presença de uma meia-lua em forma de osso do pulso especializada (do carpo semi-lunate) que permitiu que toda a mão de dobrar para trás em direção ao antebraço à maneira de moderno pássaros.
Paleopatologia
Em 2001, Ralph E. Molnar publicou uma pesquisa de patologias em terópode osso de dinossauro. Ele encontrou características patológicas em 21 gêneros de 10 famílias . Patologias foram encontrados em terópodes de todos os tamanho do corpo, embora eles eram menos comuns em fósseis de pequenos terópodes, embora isto possa ser um artefato de preservação. Eles são amplamente representada em todas as diferentes partes do terópode anatomia. Os locais mais comuns de lesão preservada e doença em dinossauro terópode do costelas e vértebras cauda. Apesar de ser abundante nas costelas e vértebras, as lesões parecem estar "ausente ... ou muito raro" em peso primário ossos de apoio dos órgãos, como o sacro, fêmur, e tíbia. A ausência de lesões conservados nestes ossos sugere que eles foram seleccionados pela evolução para a resistência à ruptura. Os locais menos conservados comuns de lesão são o crânio e dos membros anteriores, com lesões ocorrendo em cerca de igual freqüência em cada local. A maioria das patologias preservados em fósseis de terópodes são os restos de lesões como fraturas, poços, furos e, muitas vezes, provavelmente originário com mordidas. Alguns paleopathologies terópodes parecem ser evidências de infecções , que tendiam a ser confinado apenas para pequenas regiões do corpo do animal. Evidência para malformities congênitas também foram encontrados em restos terópodes. Tais descobertas podem fornecer informações úteis para a compreensão da história evolutiva dos processos de desenvolvimento biológico. Incomum fusões em elementos cranianos ou assimetrias na mesma, provavelmente, são a prova de que se está examinando os fósseis de um indivíduo extremamente antiga em vez de um doente.
História evolutiva

Durante o final do Triássico , um número de proto-terópodes e terópodes dinossauros primitivos existiu e evoluiu ao lado do outro.
O mais antigo e mais primitivo dos dinossauros terópodes eram carnívoros Eodromaeus eo herrerasaurids da Argentina (bem como, possivelmente, o onívoro Eoraptor). Os herrerasaurs existiu durante o início da tarde Triássico (Final Carniano a precoce Noriano). Eles foram encontrados na América do Norte e América do Sul e, possivelmente, também a Índia ea África do Sul. Os herrerasaurs foram caracterizados por um mosaico de características primitivas e avançadas. Alguns paleontólogos no passado considerados os herrerasaurians a ser membros de Theropoda, enquanto outros teorizou que o grupo seja saurischians basais, e pode até mesmo ter evoluído antes da divisão saurischian-ornithischian. Análise cladística na sequência da descoberta de Tawa, outro dinossauro Triássico, sugere as herrerasaurs prováveis foram os primeiros terópodes.
Os mais antigos e primitivos terópodes inequívocas (ou em alternativa, "Eutheropoda" - "verdadeiro Theropods ') são o Coelophysoidea. O coelophysoidea eram um grupo de animais amplamente distribuídas, ligeiramente construídas e potencialmente gregários. Eles incluíram pequenos caçadores como Coelophysis e predadores (possivelmente) maiores, como Dilophosaurus . Estes animais sucesso continuado do Carniano tarde (início Triássico Superior) até o Toarciano (tarde Jurássico Inferior). Embora no início classificações cladísticas eles foram incluídos no âmbito do Ceratosauria e considerado um side-ramo de terópodes mais avançados, eles podem ter sido o ancestral de todos os outros terópodes (que lhes faria um grupo paraphyletic).
A um tanto mais avançada ceratosaurs (incluindo Ceratosaurus e Carnotaurus) apareceu durante o Jurássico Inferior e continuou até o Jurássico Superior em Laurásia. Eles competiram ao lado de seus parentes mais anatomicamente avançada tetanuran e na forma de o abelisaur linhagem durou até o fim do Cretáceo em Gondwana .
O Tetanurae são mais especializado do que os ceratosaurs novamente. Eles são subdivididos em basal Spinosauroidea (alternadamente Spinosauroidea) e o derivado mais Avetheropoda. Megalosauridae eram principalmente Jurássico Médio para precoce predadores do Cretáceo, e sua restos mortais de parentes spinosaurid são na sua maioria a partir de rochas do Cretáceo início e meio. Avetheropoda, como seu nome indica, foram mais estreitamente relacionados aos pássaros e são novamente divididos em o Allosauroidea (a diversificada carcharodontosaurs) eo Coelurosauria (um grupo de dinossauros muito grande e diversificada, incluindo as aves).
Assim, durante o Jurássico tardio, não foram menos de quatro linhagens distintas de terópodes-ceratosaurs, megalosaurs, alossauros e coelurosaurs-predando a abundância de pequenas e grandes dinossauros herbívoros. Todos os quatro grupos sobreviveram até o Cretáceo, e três de ceratosaurs, coelurosaurs e alossauros-sobrevivido ao final do período, onde estavam geograficamente separada a quem-os ceratosaurs e alossauros em Gondwana, e os coelurosaurs em Laurásia.
De todos os grupos terópodes, os coelurosaurs foram de longe as mais diversas. Alguns grupos coelurosaur que floresceram durante o Cretáceo foram o tyrannosaurids (incluindo Tyrannosaurus ) a dromeossaurídeos (incluindo Velociraptor e Deinonychus , que são notavelmente similares em forma para o mais antigo pássaro conhecido, o Archaeopteryx ), o pássaro-like troodontídeos e oviraptorosaurs, o ornithomimosaurs (ou "dinossauros avestruz"), o estranho gigante-agarrada herbívoros therizinosaurs, eo avialans, que incluem aves modernas e é a única linhagem de dinossauros a sobreviver ao Cretáceo-Paleogene evento de extinção. Enquanto as raízes desses vários grupos são encontrados no Jurássico Médio, eles só se tornou abundante durante o Cretáceo adiantados. Algumas, tais como paleontólogos Gregory S. Paul, sugeriram que alguns ou todos esses terópodes avançados foram, na verdade, descendentes de dinossauros voadores ou pássaros-proto como Archaeopteryx que perderam a capacidade de voar e devolvidos a um habitat terrestre.
Classificação
História da classificação

O nome Theropoda (que significa "pés animal") foi cunhado pela primeira vez por OC Marsh em 1881. Marsh inicialmente nomeado como um Theropoda subordem para incluir o família Allosauridae, mas mais tarde expandiu o seu âmbito, re-ranking-o como uma a fim de incluir uma grande variedade de "carnívoras" famílias de dinossauros, incluindo Megalosauridae, Compsognathidae, Ornithomimidae, Plateosauridae e Anchisauridae (agora conhecido por ser herbívora prossaurópodes) e Hallopodidae (agora conhecido por ser parentes dos crocodilianos). Devido ao âmbito da Marsh Ordem Theropoda, ele veio para substituir um grupo taxonômico anterior que rival da Marsh ED Cope havia criado em 1866 para os dinossauros carnívoros, Goniopoda ("pés em ângulo").
Até o início do século 20, alguns paleontólogos, como Friedrich von Huene, deixou de ser considerado dinossauros carnívoros ter formado um grupo natural. Huene abandonado o nome Theropoda, em vez de usar Pedido de Harry Seeley Saurischia , que Huene dividido nas subordens Coelurosauria e Pachypodosauria . Huene colocado a maioria dos terópodes pequenos grupos em Coelurosauria, e os grandes terópodes e prossaurópodes em Pachypodosauria, que ele considerava ancestral ao Saurópode (prossaurópodes foram ainda pensado como carnívoro, neste momento, devido à associação incorreta de crânios rauisuchian e dentes com corpos prossaurópode, em animais, tais como Teratosaurus). Em Mateus e WD 1922 descrição de Barnum Brown do primeiro conhecido dromaeosaurid ( Dromeossauro albertensis), eles se tornaram os primeiros paleontólogos para excluir prossaurópodes dos dinossauros carnívoros, e tentaram reviver o nome Goniopoda para esse grupo, embora nenhuma dessas sugestões foram aceitas por outros cientistas.

Não foi até 1956 que Theropoda voltou para o uso como um taxon contendo os dinossauros carnívoros e seus descendentes, quando Alfred Romer re-classificados da Ordem Saurischia em duas subordens, Theropoda e Sauropoda. Esta divisão de base tenha sobrevivido em paleontologia moderna, com a excepção de, novamente, a Prosauropoda, que Romer incluído como um infraorder de terópodes. Romer também manteve uma divisão entre Coelurosauria e Carnosauria (que ele também classificou como infraordens). Esta dicotomia ficou chateado com a descoberta de Deinonychus e Deinocheirus mirificus em 1969, nenhuma das quais poderia ser classificado facilmente como "carnossauros" ou "coelurosaurs." À luz destas e de outras descobertas, no final da década de 1970 Rinchen Barsbold criou uma nova série de infraordens terópodes: Coelurosauria, Deinonychosauria, Oviraptorosauria, carnosauria, Ornithomimosauria, e Deinocheirosauria.
Com o advento de cladística e nomenclatura filogenética na década de 1980, e seu desenvolvimento nos anos 1990 e 2000, uma imagem mais clara das relações terópodes começaram a surgir. Vários grandes grupos terópodes foram nomeados por Jacques Gauthier em 1986, incluindo o clade Tetanurae por uma filial de um terópode básica dividir com outro grupo, o Ceratosauria. Como obter mais informações sobre a ligação entre dinossauros e aves veio à tona, os terópodes mais pássaro-como foram agrupadas no clado Maniraptora (também chamado por Gauthier em 1986). Estes novos desenvolvimentos também veio com um reconhecimento entre a maioria dos cientistas de que as aves surgiram diretamente dos terópodes maniraptoran e, com o abandono de postos na classificação cladística, a reavaliação de pássaros como um subconjunto de dinossauros terópodes que aconteceu para ter sobrevivido as extinções em Mesozóico o presente.
Grandes grupos



O seguinte é uma classificação simplificada de grupos terópodes com base em suas relações evolutivas, e organizado com base na lista de espécies de dinossauros do Mesozóico fornecidos por Holtz (2008). Uma versão mais detalhada pode ser encontrada em Classificação do dinossauro. A cruz (†) é usado para significar grupos sem membros vivos.
- † Herrerasauria (início dos carnívoros bípedes)
- † Coelophysoidea (pequenas, primeiros terópodes; inclui Coelophysis e parentes próximos)
- † Dilophosauridae (início com crista e terópodes carnívoros)
- † Ceratosauria (geralmente elaborada com chifres, os carnívoros sul dominantes do Cretáceo)
- Tetanurae ("caudas rígidas"; inclui a maioria dos terópodes)
- † Spinosauroidea (grupo precoce de grandes carnívoros, incluindo os espinossaurídeos semi-aquáticos)
- † Carnosauria ( Allosaurus e parentes próximos, como Carcharodontosaurus)
- Coelurosauria (penas terópodes, com uma gama de tamanhos de corpo e nichos)
- † Compsognathidae (coelurosaurs início comuns com membros anteriores reduzidas)
- † Tyrannosauridae ( Tiranossauro e parentes próximos; tinha reduzido membros anteriores)
- † Ornithomimosauria (" avestruz "; -mimics principalmente serrar; carnívoros para possíveis herbívoros)
- † Alvarezsauroidea (pequenos insetívoros com membros anteriores reduzidas cada rolamento uma garra alargada)
- Maniraptora ("ladrões de mão"; tinha, braços longos e finos e dedos)
- † Therizinosauria (herbívoros bípedes com grandes garras de mão e cabeça pequena)
- † Oviraptorosauria (principalmente desdentado; sua dieta e estilo de vida são incertos)
- † Archaeopterygidae (pequeno, terópodes alados ou pássaros primitivos)
- † Deinonychosauria (pequenas e médias empresas; pássaro-like, com uma garra toe diferenciado)
- Avialae (aves modernas e parentes extintos)
- † Scansoriopterygidae (pequenas avialans primitivos com longos dedos terceiros)
- † Omnivoropterygidae (grande, primeiros avialans de cauda curta)
- † Confuciusornithidae (pequenas avialans serrar)
- † Enantiornithes (primitivo árvore-moradia, avialans voadores)
- Euornithes (aves voadoras avançado)
- † Yanornithiformes (dentada aves chinesas Cretáceo)
- † Hesperornithiformes (aves aquáticas de mergulho especializados)
- Aves (aves modernas,-de-bico e seus parentes extintos)
Relacionamentos
A seguir cladogram é adaptado de Weishampel et al., 2004. Ele retém Coelophysoidea possível ceratosaurs, ao contrário de muitos estudos recentes colocando-os fora ceratosaurs e ancestral tanto ceratosaurs e tetanurae.
| Theropoda |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
